【CC讲坛】张蔚:蝶影重重——揭秘丛林伪装者
时间:2022-10-23张蔚
北京大学生态研究中心
北京大学生命科学学院 研究员
伪装保护,是大自然赋予多种动物的天然特性,这一物种演化中形成的机制,一直是人类探究的待解之谜。她以非模式生物枯叶蛱蝶为样本,能揭开物种从宏观到微观角度,双重演化的秘密吗?
演讲实录:
大家看到自然界里存在着各种各样的动物伪装者。

伪装者神秘低调而又生机勃勃,它们身上到底隐藏着自然赋予的什么秘密?

拟叶伪装,是一种广泛存在的动物模拟植物的现象,只要用心观察,你就能发现这些拟叶动物的踪迹,有一种因为拟叶绝技久负盛名,它就是枯叶蛱蝶。

我们实验室主要的研究方向是演化生物学,研究的是非模式生物。
而模式生物,用于揭示生命现象的普遍规律,例如果蝇、拟南芥、小鼠、线虫等等,它们研究基础比较好。

然而它们是自然界的沧海一粟,更广泛存在的其实是非模式生物,具有很多模式生物不具备的一些特征,可以用来研究更多的科学问题。
一般来讲,研究非模式生物相对困难,但是现在科学技术的发展,解锁了我们深入研究它们的一个可能性,甚至有望将它们建立成研究体系,因此这个研究过程必将充满挑战,但是也是我们探索未知的一个必经之路。

我的研究主要关注的就是蝴蝶,关注它们的蝶翅。
蝶翅的花纹是多种多样的,它们的结构相对简单,但是它的功能又很复杂,包括有运动、热调控、求偶、御敌等等。
这样一类结构简单、功能复杂的器官,我们用它来研究一些很有意思的科学问题,例如多样性如何产生、大自然又是如何塑造这些蝶翅的?
首先随着我的合作者,西藏的昆虫学家达娃老师的一个镜头,请大家从一个天敌的视角仔细观察,是否已经发现了猎物?
达娃 拍摄
当镜头逐渐拉近,是否又发现了这些蝴蝶拟叶翅花纹的区别?没有哪两片叶子是一模一样的,它展现了多样的叶形翅花纹。
刚才视频里展示的,是枯叶蛱蝶的翅腹面,它们闭合着翅,但是当它们飞翔的时候,就会展示它的翅背面,带有鲜艳的斑块。

其实早在1867年,它们就被演化生物学的先驱华莱士所发现,并写在了书中。
2015年,我在芝加哥大学生态与演化系做博士后研究,有一天,我在读华莱士的这本书,给我留下了深刻的印象。

枯叶蛱蝶在我国也属于“三有动物”(有益、有重要经济价值、有科学研究价值),以保护性的伪装而出名。
这类蝴蝶的蝶翅有背、腹面迥异的花纹,显示出它可能受到不同的选择压力。
这些假设,还有待于我们进一步地去研究和证明,因此我们这个枯叶蛱蝶的体系,可以用来研究不同选择压力下驱动的表型多样性,此外还可以用来研究背腹面表型不对称的发育机制。

我们以枯叶蛱蝶的蝶翅为模式研究生物多样性,可以研究包括遗传多样性、物种多样性,以及受到环境因素和遗传因素影响的表型多样性,我就意识到这是一个很好的研究体系。

要研究枯叶蛱蝶,首先还要了解它的生活史。它从受精卵孵化,在幼虫期要经历多次蜕皮,然后结蛹,破茧成蝶,这就是一种完全变态发育的昆虫。
在幼虫期,蝴蝶离不开赖以为食的寄主植物。因此我们要做蝴蝶的研究,还要去在实验室饲养它们的寄主植物,给它们做口粮,这对于非模式生物的研究来说,也是一个大工程。
大家可以猜一下,我们把一只枯叶蛱蝶养大成蝶,需要多少寄主植物?需要十五盆,这也是研究非模式生物的一个特色。
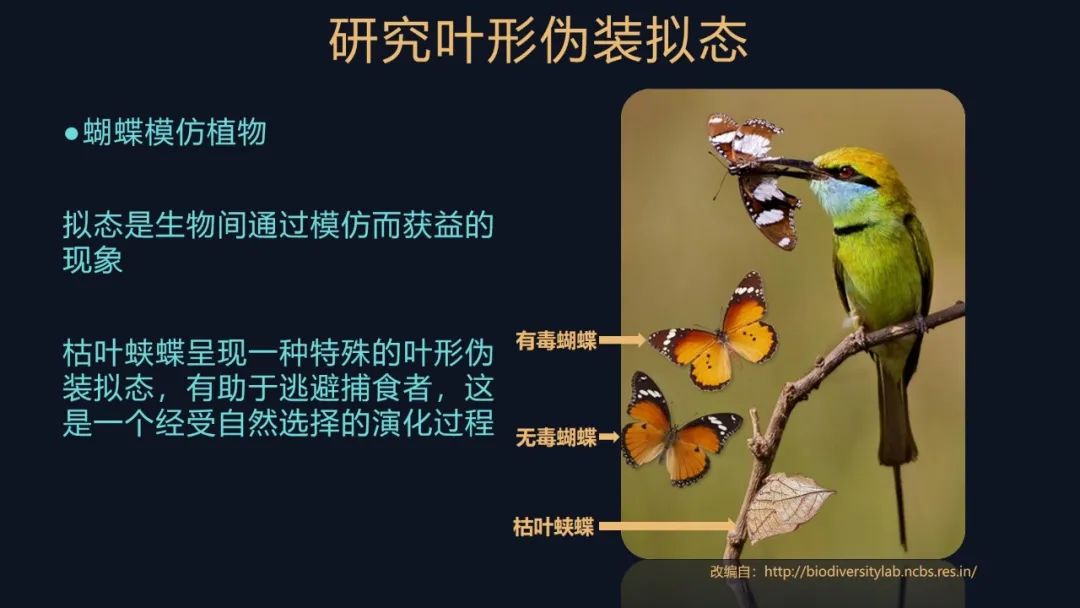
我们今天要介绍的是蝴蝶模仿植物,我们称之为叶形拟态。
总而言之,拟态的蝴蝶就逃避了捕食者,是有好处的。它们展现的这些巧妙的拟态,其实是经受了大自然的选择的产物,这是一个被动选择的演化过程。

生物演化可以追溯至生命起源的一个阶段,我们都共享一棵生命之树,它枝繁叶茂,它也不是一种简单的线性的演替,让我们看到的这个世界多姿多彩,这是生物多样性的一个魅力所在。

演化本身是没有方向的,也没有低级和高级之分,但是选择压力,在一定的时间可能有一定的方向。
只有选择压力本身,没有可供选择的材料也不行,所以我们的遗传信息里,还会存在一定的变异,这些都是生物演化领域关注的问题。
有了这些研究的框架和手段,我们就做好准备来揭开今天主角的一个神秘面纱。
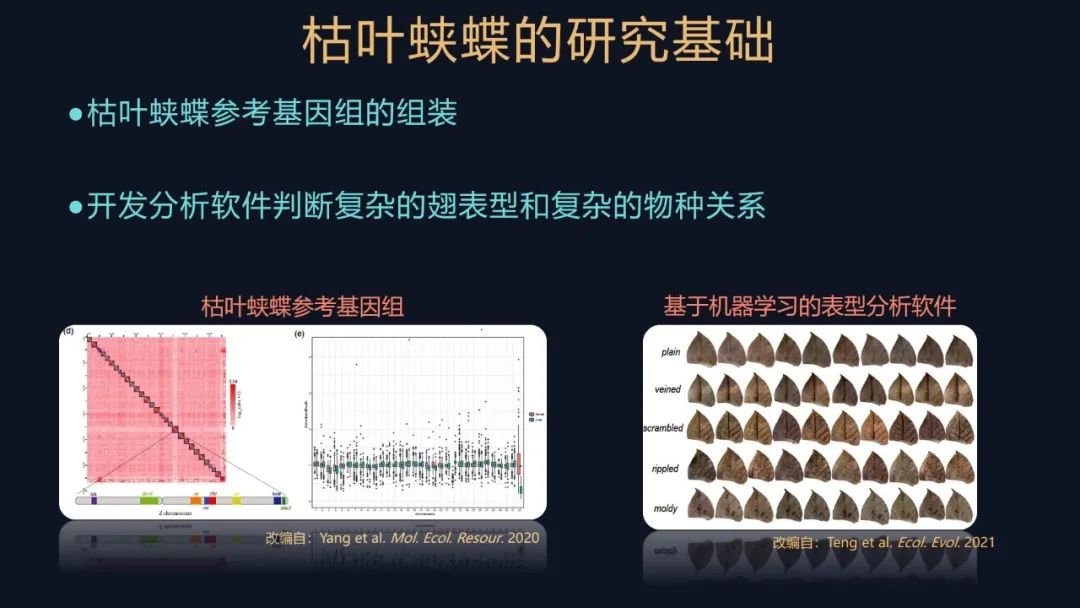
我们首先要参与组装枯叶蛱蝶中华亚种的参考基因组,我们还实现了在实验室条件下饲养这些枯叶蛱蝶和开展实验,此外我们还开发了一个基于深度学习的软件,来分析它们复杂的物种关系。
有了这些研究基础,我们就需要采集样本了。枯叶蛱蝶在分类学中是一个属,分布的范围很广泛,还包括多个物种。要弄清楚样品所属的物种和它的关系,也是我们开展研究的一个重要的前提。

首先我们从宏观演化的水平,来解析枯叶蛱蝶属的物种多样性,解析其物种的起源和演化。我们就要去到它们自然的生境里,对它们进行观察,足迹遍布典型的分布的区域。我们采集枯叶蛱蝶的地区,大部分都是热带和亚热带的雨林,例如有西藏、云南、海南等等。
我们幸运地躲过了毒蛇,但是很多人没有躲过蚂蝗,没有躲过“一点红”的追捕,我们就有博士后曾经被“一点红”咬了手,手就肿得像个馒头一样,但还是坚持每天在收集采集工作。

在这个过程里最令我印象深刻的一件事,是我们习惯用捕虫网对它们(蝴蝶)进行采集,有时候难以追上它们隐蔽而迅速的身影,后来我们根据枯叶蛱蝶成虫喜食一些发酵水果汁液的习性,准备了诱笼,并为它们奉上混合着啤酒和西瓜的一些大餐(我们认为是大餐),但是收效甚微,有时候遇到降雨,把食物冲散了,有时候即使食物完好也收效甚微,吸引不到它们,最后我们恍然大悟,其实雨林里并不缺乏这些腐烂的植物果实,我们准备的大餐显然不够有吸引力。
我们的同学就灵机一动,去当地的一个菜市场买了点啥呢?买了鸡肠、鸡杂,经过阳光暴晒,可想而知要怎么样!它发出刺鼻的味道,让人避之尤恐不及。但是就是这种味道,特别受蝴蝶的青睐,纷纷现身大快朵颐。
这就太好了!我们终于找到了枯叶蛱蝶的一个大餐,虽然人都被熏得受不了,但是纷纷地表示这个办法太绝妙了!
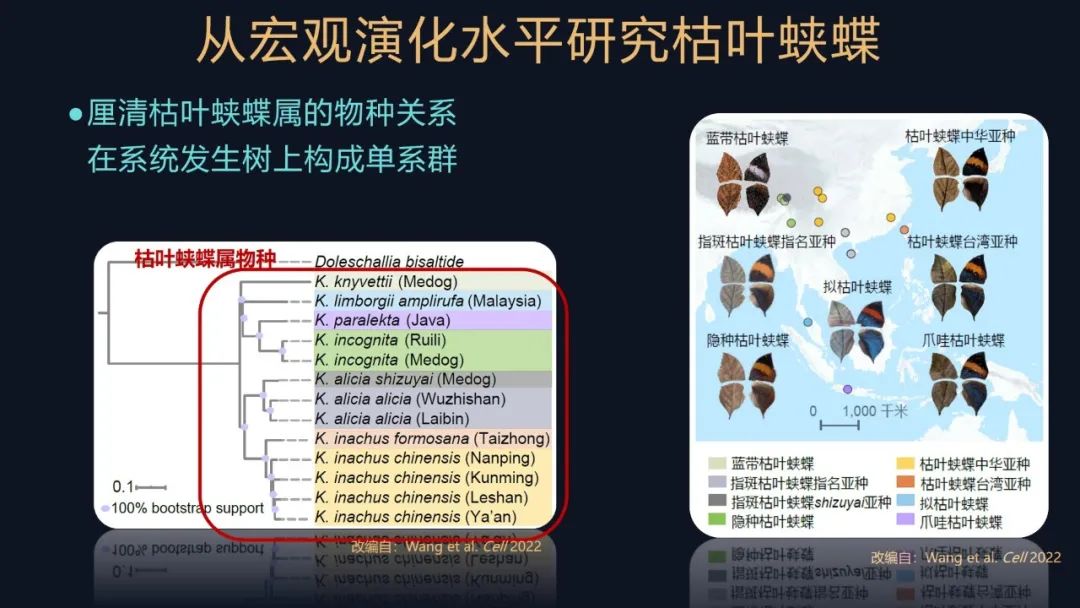
很幸运的是,我们还得到了一些罕见的样本。这个图上所示的,就是一些枯叶蛱蝶的,我们主要的采集区域。
我们还厘清了这些枯叶蛱蝶的物种关系,发现它们在系统发生树上都构成一个单系群,这些也都是我们后续开展研究的一个基础。
此外为了研究这些叶形拟态的蝴蝶的起源,我们还收集了很多的蛱蝶科的样本,包括有一些叶形拟态的其他的蝶,例如有蠹叶蛱蝶、美洲枯叶蝶等等,听听它们的名字都跟叶形、拟叶形有关。
但是就是在这个系统发生树里,这些标出了绿色的样本,可以看出它们构成了一个多系群。
枯叶蛱蝶属的样品是广泛存在的,一般在一个地理分布区,只有一个物种或者亚种,我们能采到这些样品。

但是在一个地方,就是我刚才提到的西藏的墨脱县,我们获得了多个物种的样本,一开始我们以为这是不是弄错了,没料到这是个发现,等到我们把样品的这些基因组全都分析、验证了以后,我们才相信了这个事实,这就奇怪了,这里为什么会有这么多的枯叶蛱蝶的物种呢?
我们因此就提出了假设,认为这个区域可能是枯叶蛱蝶属的一个分化中心,或者也有可能是它们在历史上的冰期的一个避难所?
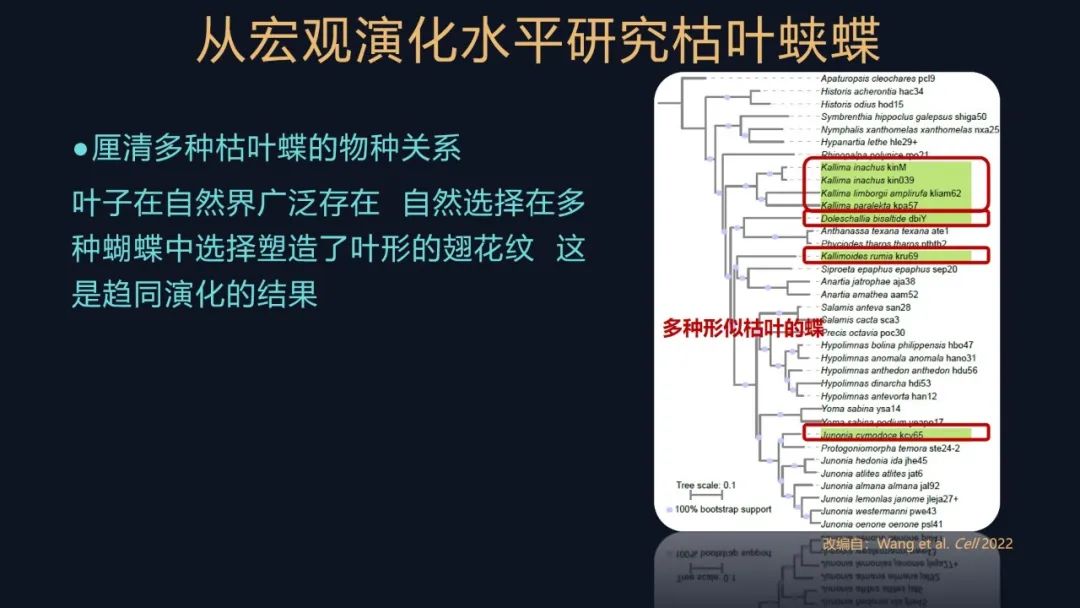
自然界的叶子广泛存在,自然选择也多次选择了这些叶形的表型,这些叶形的翅花纹,就是在蛱蝶科多次独立的起源,它属于一种趋同演化。
这样的话,我们不仅把枯叶蛱蝶属的物种关系厘清了,我们还把其他拟叶型的蛱蝶科物种的关系都弄明白了,这让我们感到很开心。
根据我们刚才提出的假设——关于枯叶蛱蝶为什么在墨脱这个地方有多种物种存在,我们就进一步地研究蛱蝶属的起源和分化了。
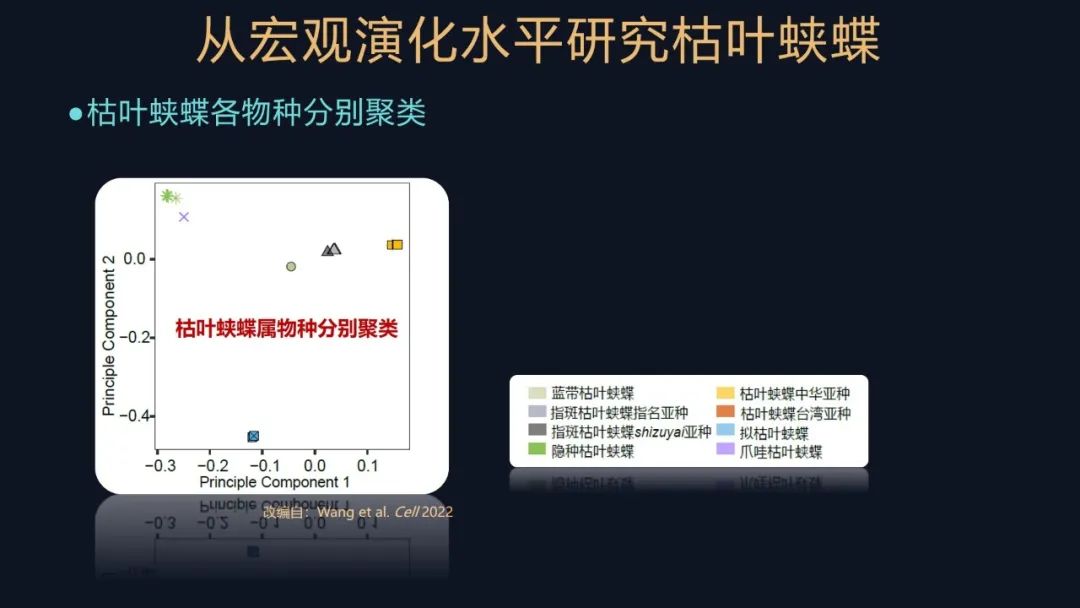
我们对它们的遗传信息进行聚类和研究,发现这些物种或者亚种,都根据它们的亲缘关系聚类了,这说明我们之前对它们的研究和分类还是比较合理和可靠的。
我们又研究了这些蛱蝶物种在历史上的种群大小的一个变化,这张图其实就是根据它们的遗传信息,反映从现在到以前,这些不同物种种群的一个变化情况。可以看到大部分的枯叶蛱蝶属的物种或亚种,有比较大的种群,但是都是在末次冰期这种历史时期,它经历了一个瓶颈效应的影响,也就是它的种群有收缩、有波动,但是总体来说在喜马拉雅东部地区的这些物种,都显示出了比较大的种群,再次说明这个区域对于枯叶蛱蝶属是比较重要的。
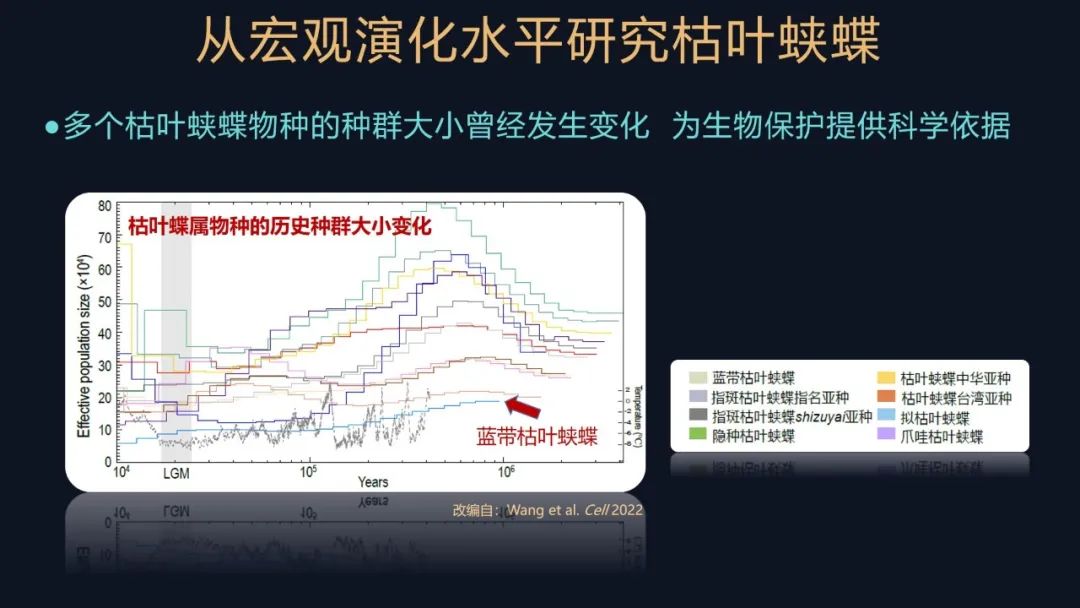
还有一个例外,就是这张图上的蓝带枯叶蛱蝶,它始终维持着一个比较小的种群,其实这个蝴蝶目前在我们国家,仅仅在墨脱县发现分布了,它的样品非常难以获得,我们大概要采集几年才能获得这样一个样本,再次印证了它的种群比较小的这一点。
我们其实做昆虫研究,很难像研究大型的动物一样,对这些昆虫进行抽样计数,你仔细地把它数一数,这是不可能的,但是我们通过基因组学的研究手段,可以获得它们这些种群的信息,这也为对它们进行生物保护提供了很重要的科学依据。
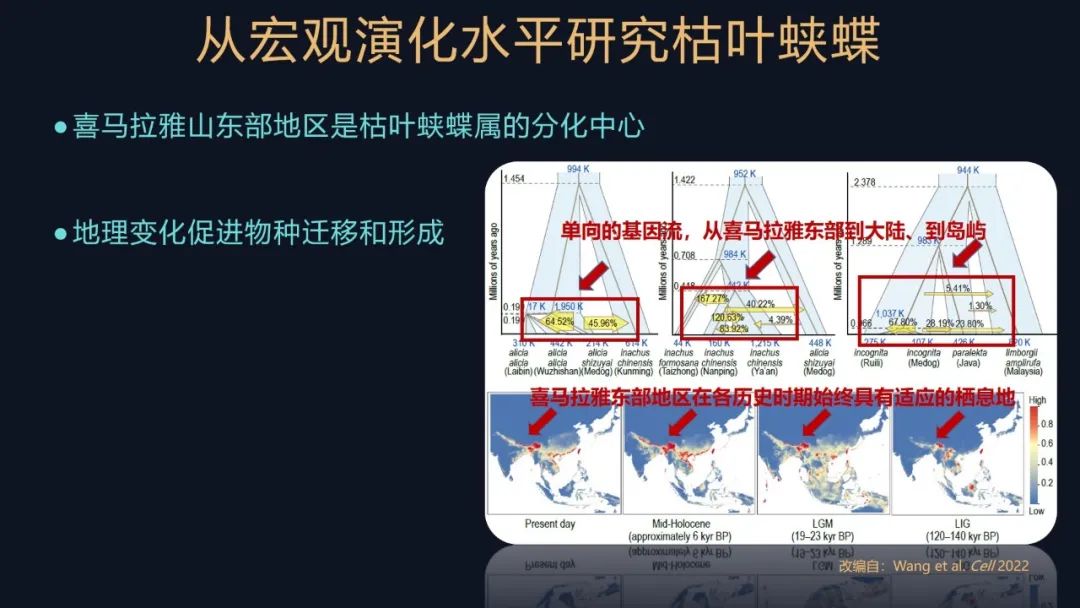
我们还进行了一些基于模型的分析,研究不同的亚种、或者是不同的种群之间的基因流。所谓基因流,就是指遗传信息的一个交换。
我们这个研究就发现了枯叶蛱蝶属都有一些单向的基因流,方向是什么样的呢?例如从喜马拉雅东部地区向大陆地区流动,或者是从大陆地区到岛屿地区流动,反映了枯叶蛱蝶的一个迁移和分化的过程。这个过程是需要非常长的历史时间的,非常壮丽的一个迁移的过程。
我们还对它们历史上适宜的栖息地进行分析,发现喜马拉雅东部区域始终有它们适宜的一个生境,而冰期由于海平面下降,会产生一些陆桥的结构,又会帮助这些蝴蝶向岛屿进行进发,进而分化形成了一些岛屿的物种,这就是一幅波澜壮阔的迁移图景,持续数十万年。
枯叶蛱蝶属的分化时间,和青藏高原隆起的时间是相符合的。高原隆起产生的海拔落差,会形成多样的生境,它有助于孕育新的枯叶蛱蝶的物种,到现在为止我们就揭示了枯叶蛱蝶属的演化和起源的历史。
它们的叶形翅型又如何产生呢?我们将从微观演化的水平解析它们叶形表型的一些遗传机制。

我们先以其中的一种,枯叶蛱蝶的中华亚种作为一个模式,对它们进行观察,建立一些家系。我们把具有不同叶形表型的这些个体进行组合,看它后代会有怎样的表型。
在这个过程里,我们发现着实不少,它一共有十种离散的表型。

有比较朴素,没有花纹的,我们命名为无主脉表型;还有的是有一条深色的主脉,我们命名为有主脉;有的还有侧脉,有分支了;还有的有霉斑、有水波纹;不同的表型还会有一定的规律进行组合。这其实是非常经典的遗传学的研究方法,只不过我们把它用在了非模式的生物上了。
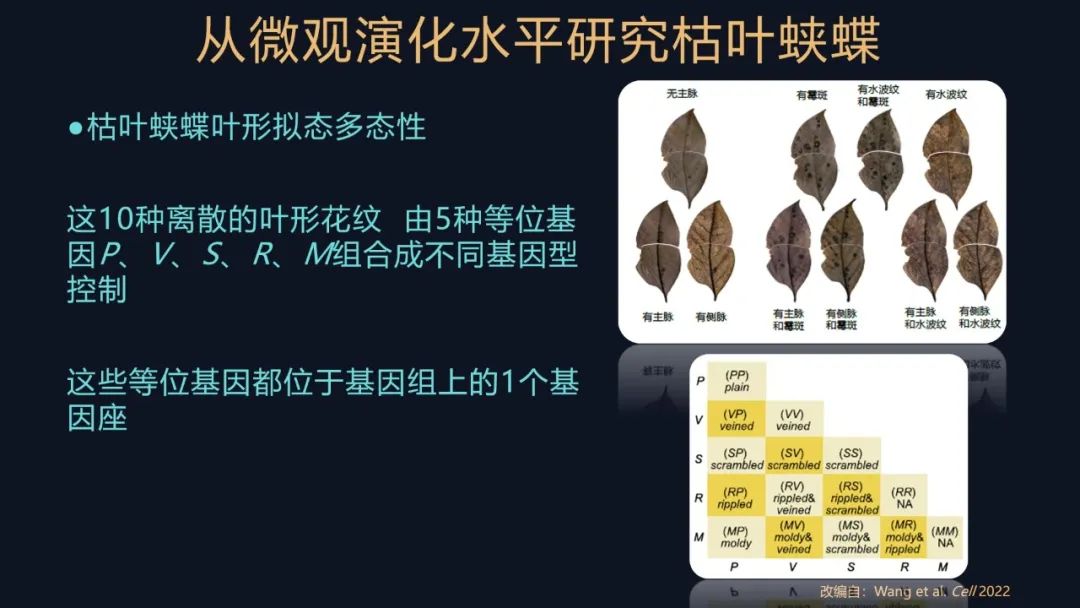
我们还对这些表型进行一个判断,推测这十种表型,由五种等位基因控制,都位于一个单基因座,它们通过组合形成不同的基因型,进而控制这十种表型,但这些都是基于我们初步的实验结果提出的假设,我们还要对它进行验证。
但是逐渐我们的实验脉络就愈加清晰了,发现了这么多的叶形表型 非常地惊喜,但是也意味着整个的研究过程比我们预期的还要复杂。
但科学研究中最有趣的事儿就是在此,莫过于你提出一个或回答一个问题的过程里,陆续发现更多有意思的和待解答的问题。
我们用饲养获得的个体,去根据它们的不同的表型进行分组,然后对它们进行全基因组的关联分析,解析控制叶形拟态的遗传机制。
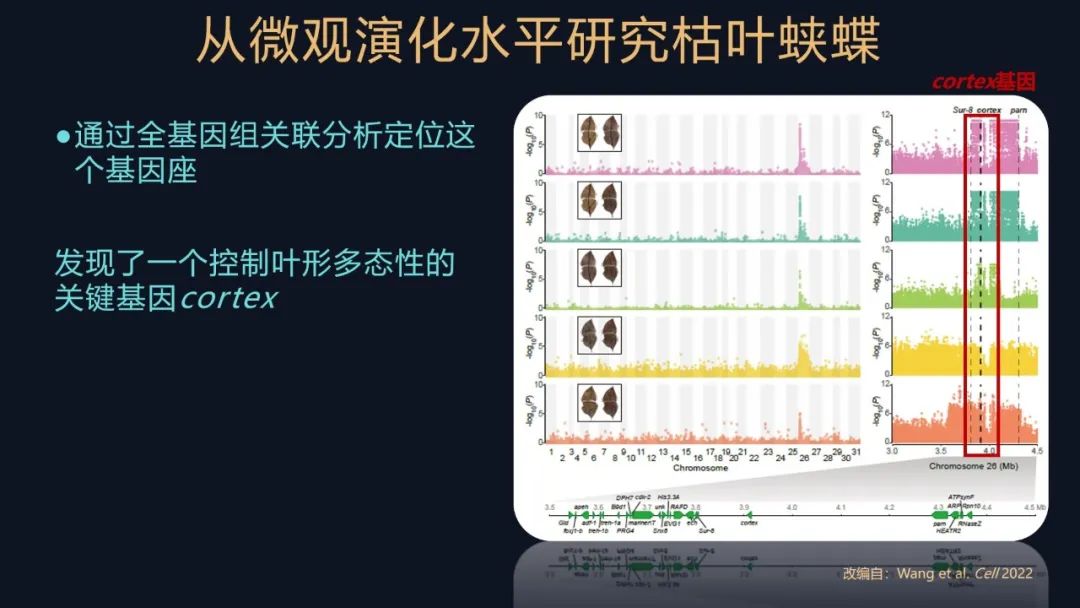
我们把两两表型进行一个分析比较,都在基因组的同一个位置发现了类似的信号,这个信号里有啥呢?
我们发现了一个控制拟态的关键基因,它叫做cortex 。
这个发现又是一个意外之喜,为什么这么说?cortex这个基因,它是一个大名鼎鼎的基因。蝴蝶的发育过程相对来讲其实是比较保守的,有一些工具盒基因各司其职,它们负责起到一些重要的作用,cortex基因就是其中的之一。

它已经被报道参与了多种受到自然选择产生表型多样性的例子里了。

这个基因的缺少确实会使枯叶蛱蝶的翅的腹背面花纹受到影响。进一步我们还想知道一下,这个多样的叶形表型,它是如何维持的?
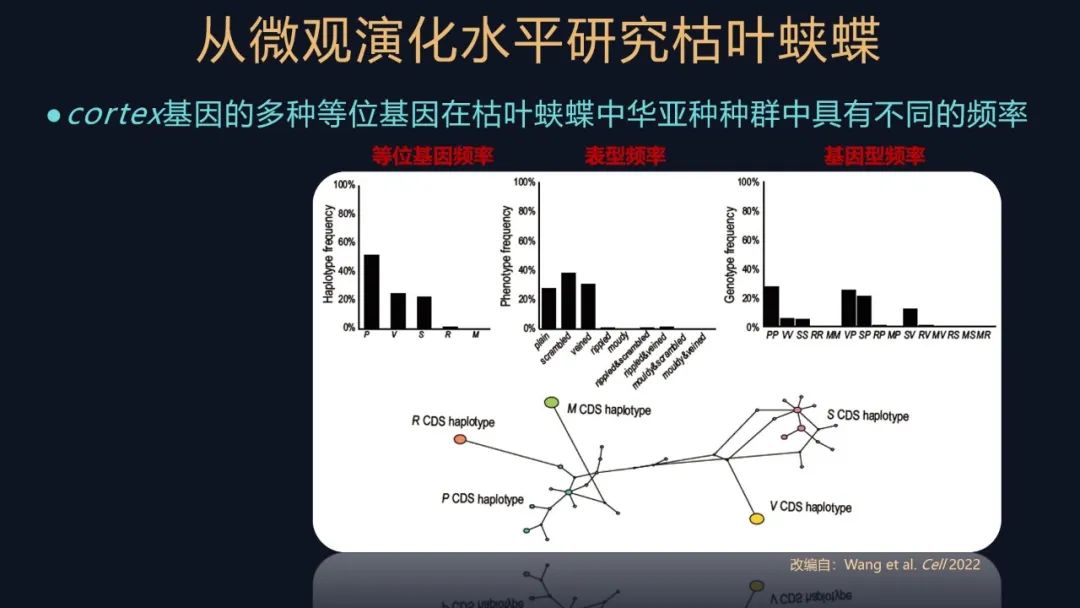
我们就去统计枯叶蛱蝶中华亚种不同的基因型和表型的一个频率,这又是一个非常经典的遗传学的研究方法。发现它们都各不相同,说明cortex不同的等位基因,它们已经演化了比较久的时间了。其实不仅仅是在枯叶蛱蝶中华亚种的种群,我们在很多的枯叶蛱蝶属的物种,都发现了叶形拟态的多态性。
这个结果让人眼前一亮,又是一个未知忽然出现的惊喜。
这可能是一个平衡选择的例子,这些等位基因产生的时间,刚好跟整个枯叶蛱蝶属的物种分化的时间是能对得上的,进而又跟一个什么时间对得上?就是跟青藏高原隆起的时间又对上了。这是一个受到自然选择维持的一个长期的平衡选择的一个例子,它很罕见。

为什么说罕见呢?要维持长期的平衡选择,对于这些物种来讲压力其实是比较大的。
我们建立了模型证明,它受到了一种负频率依赖的选择,和一种纯和的劣势。
例如在枯叶蛱蝶中华亚种里,我们发现有一些表型,它只能以杂合的形式存在,就是有一些刚刚产生出来的,例如霉斑表型,没有一个纯和的状态,这说明什么?可能它还是新产生出来的等位基因,还有一些劣势需要通过杂合状态来弥补;
而负频率依赖的选择又是什么意思?说明什么表型多了、频率多了,它反而不好。试想如果捕食者看到了整齐划一,千篇一律的叶形,比我们眼中的假花、假草还不能再假了,一眼假,很快就会被捕食者所识破。
所以它们的这种多种多样的叶形,恰恰好是自然的伪装,所以这是自然选择的一个结果,也是枯叶蛱蝶属的一个生存策略。

我们在微观和宏观演化水平研究了枯叶蛱蝶属的起源和多样化的工作,最近也以封面文章的形式发表在了《细胞》杂志上。

我们究竟从研究枯叶蛱蝶的叶形伪装里获得一些怎样的启发?
我们首先解锁了一个新的研究体系,通过它还研究了生物多样性的演化和遗传机制。
关于叶形伪装拟态的研究也都是刚刚开始,这是枯叶蛱蝶作为研究体系的一个魅力所在,也是科学研究的魅力所在。

孤举者难起,众行者易趋,当我们克服重重困难,再次回顾的时候,非模式生物的研究,它们就都变成了科学求索道路上的风景。
